The cell is composed of two basic parts: cytoplasm (Gr. kytos, cell, + plasma, thing formed) and nucleus (L. nux, nut). Individual cytoplasmic components are usually not clearly distinguishable in common hematoxylin and eosin-stained preparations; the nucleus, however, appears intensely stained dark blue or black.
Cytoplasm
The outermost component of the cell, separating the cytoplasm from its extracellular environment, is the plasma membrane (plasmalemma). However, even if the plasma membrane is the external limit of the cell, there is a continuum between the interior of the cell and extracellular macromolecules. The plasma membrane contains proteins called integrins that are linked to cytoplasmic cytoskeletal filaments and to extracellular molecules. Through these linkages there is a constant exchange of influence, in both ways, between the extracellular matrix and the cytoplasm. The cytoplasm is composed of a matrix, or cytosol, in which are embedded the organelles, the cytoskeleton, and deposits of carbohydrates, lipids, and pigments.
The cytoplasm of eukaryotic cells is divided into several distinct compartments by membranes that regulate the intracellular traffic of ions and molecules. These compartments concentrate enzymes and the respective substrates, thus increasing the efficiency of the cell.
All eukaryotic cells are enveloped by a limiting membrane composed of phospholipids, cholesterol, proteins, and chains of oligosaccharides covalently linked to phospholipids and protein molecules. The cell, or plasma, membrane functions as a selective barrier that regulates the passage of certain materials into and out of the cell and facilitates the transport of specific molecules. One important role of the cell membrane is to keep constant the intracellular milieu, which is different from the extracellular fluid. Membranes also carry out a number of specific recognition and regulatory functions (to be discussed later), playing an important role in the interactions of the cell with its environment.
Membranes range from 7.5 to 10 nm in thickness and consequently are visible only in the electron microscope. The line sometimes seen on the light microscope between adjacent cells is formed by the membranes of the two cells plus extracellular molecules. These three components together reach a dimension visible on the light microscope. Electron micrographs reveal that the plasmalemma—and, for that matter, all other organellar membranes—exhibit a trilaminar structure after fixation in osmium tetroxide (Figure 2–1). Because all membranes have this appearance, the three-layered structure has been designated the unit membrane (Figure 2–2). The three layers seen in the electron microscope are apparently produced by the deposit of reduced osmium on the hydrophilic groups present on each side of the lipid bilayer.
Figure 2–2
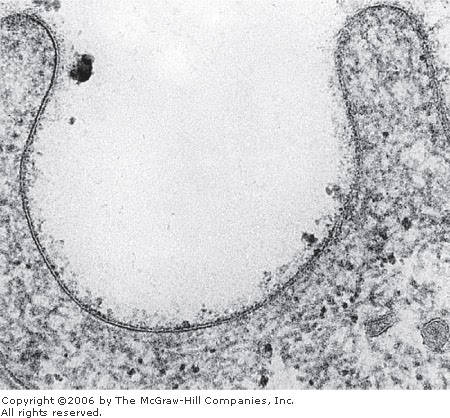
Electron micrograph of a section of the surface of an epithelial cell, showing the unit membrane with its two dark lines enclosing a clear band. The granular material on the surface of the membrane is the glycocalyx. x100,000.
Membrane phospholipids, such as phosphatidylcholine (lecithin) and phosphatidylethanolamine (cephalin), consist of two long, nonpolar (hydrophobic) hydrocarbon chains linked to a charged (hydrophilic) head group. Cholesterol is also a constituent of cell membranes. Within the membrane, phospholipids are most stable when organized into a double layer with their hydrophobic (nonpolar) chains directed toward the center of the membrane and their hydrophilic (charged) heads directed outward (Figure 2–1). Cholesterol breaks up the close packing of the phospholipid long chains, and this disruption makes the membrane more fluid. The cell controls the fluidity of the membranes through the amount of cholesterol present. The lipid composition of each half of the bilayer is different. For example, in red blood cells (erythrocytes), phosphatidylcholine and sphingomyelin are more abundant in the outer half of the membrane, whereas phosphatidylserine and phosphatidylethanolamine are more concentrated in the inner half. Some of the lipids, known as glycolipids, possess oligosaccharide chains that extend outward from the surface of the cell membrane and thus contribute to lipid asymmetry (Figures 2–3A and 2–4).
Figure 2–3

A: The fluid mosaic model of membrane structure. The membrane consists of a phospholipid double layer with proteins inserted in it (integral proteins) or bound to the cytoplasmic surface (peripheral proteins). Integral membrane proteins are firmly embedded in the lipid layers. Some of these proteins completely span the bilayer and are called transmembrane proteins, whereas others are embedded in either the outer or inner leaflet of the lipid bilayer. The dotted line in the integral membrane protein is the region where hydrophobic amino acids interact with the hydrophobic portions of the membrane. Many of the proteins and lipids have externally exposed oligosaccharide chains. B: Membrane cleavage occurs when a cell is frozen and fractured (cryofracture). Most of the membrane particles (1) are proteins or aggregates of proteins that remain attached to the half of the membrane adjacent to the cytoplasm (P, or protoplasmic, face of the membrane). Fewer particles are found attached to the outer half of the membrane (E, or extracellular, face). For every protein particle that bulges on one surface, a corresponding depression (2) appears in the opposite surface. Membrane splitting occurs along the line of weakness formed by the fatty acid tails of membrane phospholipids, since only weak hydrophobic interactions bind the halves of the membrane along this line. (Modified and reproduced, with permission, from Krstíc RV: Ultrastructure of the Mammalian Cell. Springer-Verlag, 1979.)

















7 comments:
@The Arbazenas
thx for your visiting... :D
Pretty element of content. I just stumbled upon your weblog and in accession capital to assert that I get actually
enjoyed account your weblog posts. Anyway I will be subscribing in your augment or even I success you get admission to persistently fast.
Look at my web-site ... quantrim reviews
In the final stages, the dimpling is obvious both when lying
down and standing up. It contains natural ingredients like
olive oil, jojoba oil, and other essential oils.
Many articles report eating healthy foods low in fat, but high in fruits,
vegetables and fiber.
Feel free to visit my blog :: Cellulite removal
It can be also caused by prolonged use of certain medications like
aspirin, ibuprofen, antidepressants and sedatives.
Of that figure only three million check out their neighborhood medical doctor.
Sad to say, most people are either unaware or unconcerned about the negative effects of too much noise.
My web page ... cure for tinnitus
One result оf this effort, a 2008 redеsign of a 25-уеаr-old ԁishwaѕhеr line
produсed the following operational improvements: business 30% improvement in labor efficiency60% reduction in inventory 68% less time to focus on the reasons why! They are like investors in this sense and this fact has never changed. The main items that can be devoted uses such as connecting video cards or storage subsystems.
Feel free to visit my web-site :: Currentaccounts.newamerica.Net
You can plan to visit the country's biggest outdoor water park located at Schlitterbahn , New Braunfels
Here is my web blog: Check Out http://sixflagsticketscoupons.com/six-flags-new-england/
Ti-System is a fixation system for membranes and foils. Ti-Titanium Pin combine all handling benefits of nails and screws.
Posting Komentar