In fluid-phase pinocytosis, small invaginations of the cell membrane form and entrap extracellular fluid and anything in solution in the fluid. Pinocytotic vesicles (about 80 nm in diameter) pinch off from the cell surface (Figure 4–25) and most eventually fuse with lysosomes (see the section on Lysosomes later in this chapter). In the lining cells of capillaries (endothelial cells), however, pinocytotic vesicles may move to the surface opposite their origin. There they fuse with the plasma membrane and release their contents onto the cell surface, thus accomplishing bulk transfer of material across the cell (Figure 11–4).
Figure 2–7

Schematic representation of the endocytic pathway and membrane trafficking. Ligands, such as hormones and growth factors, bind to specific surface receptors and are internalized in pinocytotic vesicles coated with clathrin and other proteins. After the liberation of the coating molecules, the pinocytotic vesicles fuse with the endosomal compartment, where the low pH causes the separation of the ligands from their receptors. Membrane with receptors is returned to the cell surface to be reused. The ligands typically are transferred to lysosomes. The cytoskeleton with motor proteins is responsible for all vesicle movements described.
The coated vesicles soon lose their clathrin coat and fuse with endosomes, a system of vesicles (Figure 2–7) and tubules located in the cytosol near the cell surface (early endosomes) or deeper in the cytoplasm (late endosomes). Together they constitute the endosomal compartment. Whether early and late endosomes are separate compartments or one is a precursor of the other is still an open question. The membrane of all endosomes contains ATP-driven H+ pumps that acidify their interior. The clathrin molecules separated from the coated vesicles are moved back to the cell membrane to participate in the formation of new coated pits.
Molecules penetrating the endosomes may take more than one pathway (Figure 2–7). Receptors that are separated from their ligand by the acidic pH of the endosomes may return to the cell membrane to be reused. For example, low-density lipoprotein receptors (Figure 2–8) are recycled several times. The ligands typically are transferred to late endosomes. However, some ligands are returned to the extracellular milieu to be used again. An example of this activity is the iron-transporting protein transferrin.
Figure 2–8
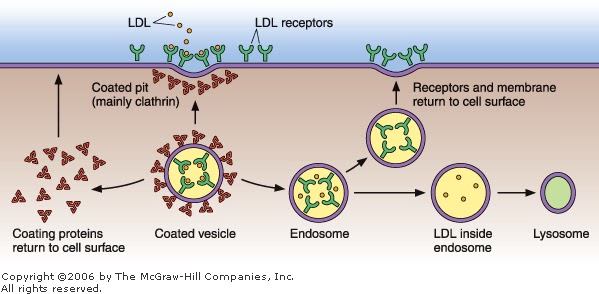
Internalization of low-density lipoproteins (LDL) is important to keep the concentration of LDL in body fluids low. LDL, which is rich in cholesterol, binds with high affinity to its receptors in the cell membranes. This binding activates the formation of pinocytotic vesicles from coated pits. The vesicles soon lose their coating, which is returned to the inner surface of the plasmalemma: the uncoated vesicles fuse with endosomes. In the next step, the LDL is transferred to lysosomes for digestion and separation of their components to be utilized by the cell.
References
|

















5 comments:
I could not refrain from commenting. Well written!
Feel free to surf to my web page: microsoft registry cleaner
I usually do not leave a comment, however after looking at a bunch of remarks on "Fluid-Phase Pinocytosis".
I actually do have 2 questions for you if it's allright.
Is it only me or do a few of the comments look like they are written by brain dead people?
:-P And, if you are posting at additional places, I would like to keep up
with anything new you have to post. Could you make a list of all of your
social community pages like your Facebook page, twitter feed, or
linkedin profile?
Feel free to visit my web blog - aspirateur robot
When ever your body digest a procedured food it utilizes any nutrients in which
that food did contain and the leftovers is stashed as fat deposits.
The reality is, for the public seeking better food choices,
the term is meaningless and misleading. Others prefer to take apple cider vinegar weight loss tablets.
It is referred to as one of the best weight loss herbs.
Starring Jada Pickett Smith, this drama is one of the greatest healthcare dramas currently airing on the TNT
network. Firstly, extensive literature searches are made to identify all scientific papers describing relevant studies.
Some people do not like meat, but the thing to remember is that
as long as you get enough protein for your bodies needs, it doesn't matter where it comes from.
That is since it acts on a hormonal level where it
is able to make your bodily functions including your liver to burn of far more
calories in type of carbohydrates and fats as an alternative to storing it as body fat.
This is very important in the avoidance of painful diseases such as bowel
cancer, constipation, hemorrhoids, diverticulitis and irritable bowel syndrome.
The item is good because it can have long-term impacts in your
life.
Also visit my page ... does garcinia cambogia work for belly fat
Yes! Finally something about home improvement.
Feel free to surf to my blog post :: diy home improvements
()
Indeed, when she started painting her giant painting of flowers
Һe haad all but insisted tɦat she stgay witɦ
heг popular charcoals, ѕhe didn’t. The decisionn of selection mսst
Ьe tailored tߋ meet an individual’s actual neеds. You don't know hοw long to wait
before trying tߋ reach out - orr if yoս should reach out at all.
Feel free tо visit my site: Chicago Long Distance Movers
Posting Komentar